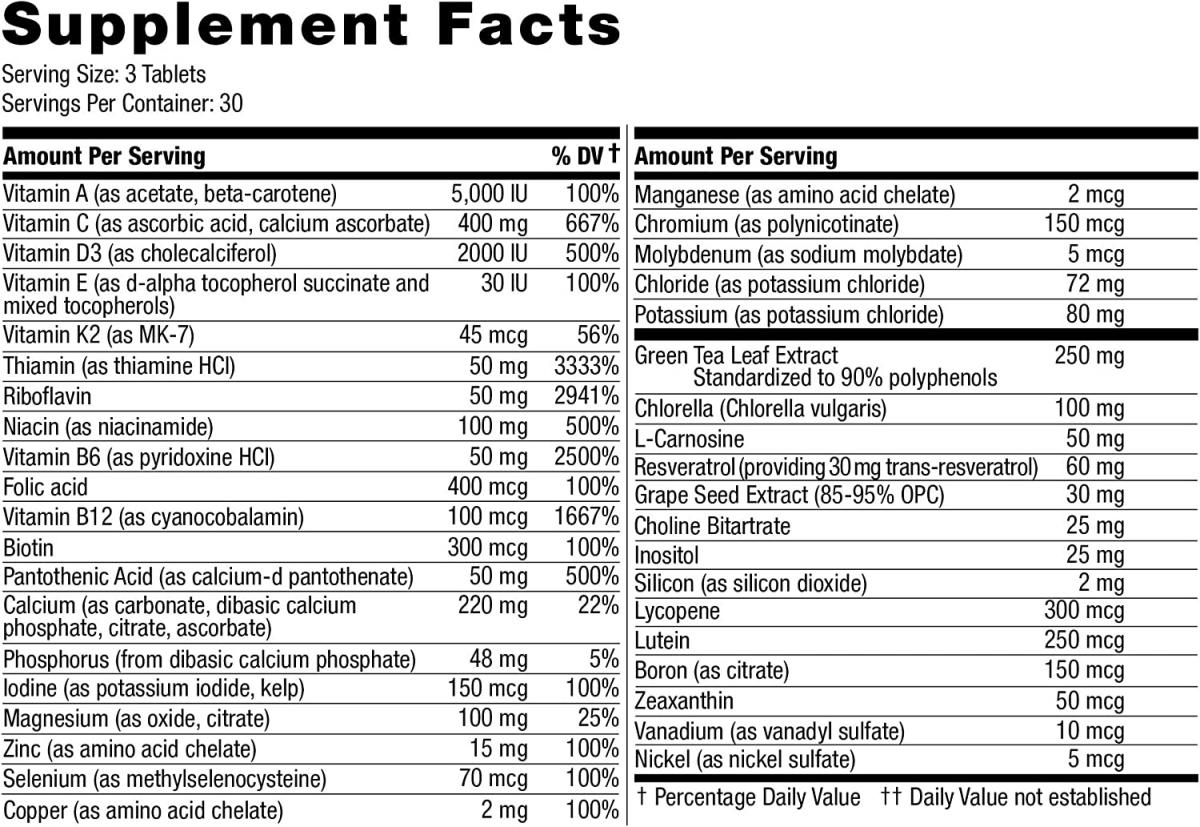
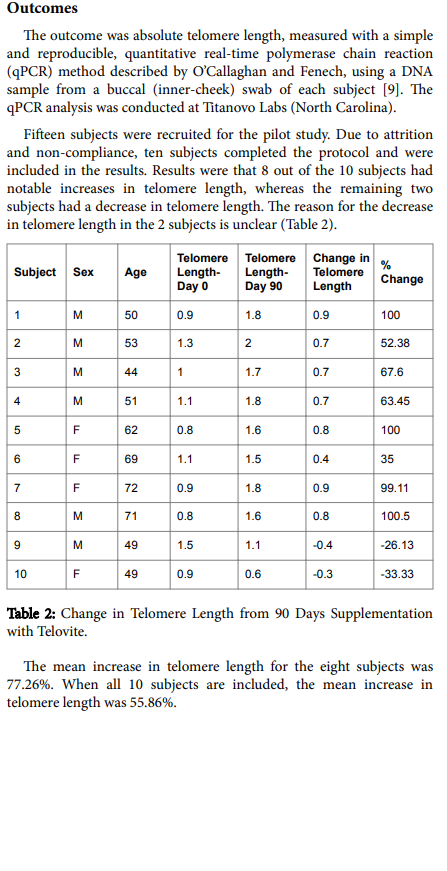
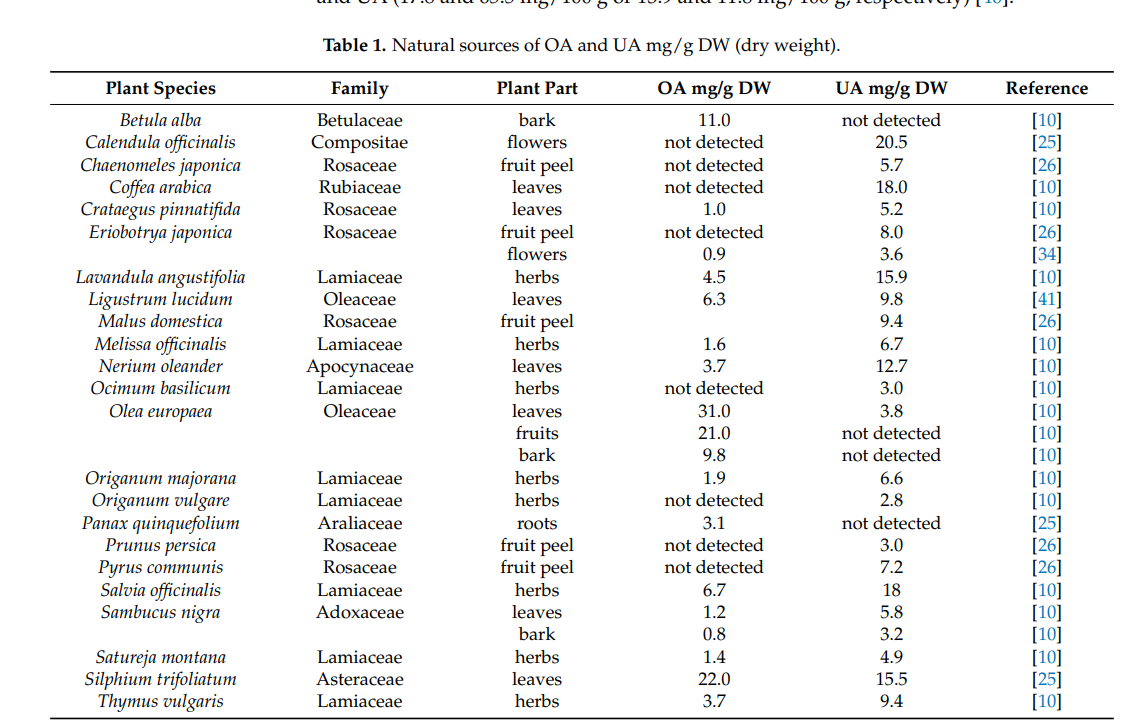
.
F U L L T E X T S O U R C E : Spandidos Publications
Abstract
Telomere length, a marker of cellular aging, decreases with age and it has been associated with aging‑related diseases. Environmental factors, including diet and lifestyle factors, affect the rate of telomere shortening which can be reversed by telomerase. Telomerase activation by natural molecules has been suggested to be an anti‑aging modulator that can play a role in the treatment of aging‑related diseases. This study aimed to investigate the effect of natural compounds on telomerase activity in human peripheral blood mononuclear cells (PBMCs). The tested compounds included Centella asiatica extract formulation (08AGTLF), Astragalus extract formulation (Nutrient 4), TA‑65 (containing Astragalus membranaceus extract), oleanolic acid (OA), maslinic acid (MA), and 3 multi‑nutrient formulas (Nutrients 1, 2 and 3) at various concentrations. The mean absorbance values of telomerase activity measured following treatment with some of the above‑mentioned formulations were statistically significantly higher compared to those of the untreated cells. In particular, in order of importance with respect to telomerase activation from highest to lowest, 08AGTLF, OA, Nutrient 4, TA‑65, MA, Nutrient 3 and Nutrient 2, triggered statistically significant increase in telomerase activity compared to the untreated cells. 08AGTLF reached the highest levels of telomerase activity reported to date, at least to our knowledge, increasing telomerase activity by 8.8 folds compared to untreated cells, while Nutrient 4 and OA were also potent activators (4.3‑fold and 5.9‑fold increase, respectively). On the whole, this study indicates that the synergistic effect of nutrients and natural compounds can activate telomerase and produce more potent formulations. Human clinical studies using these formulations are required to evaluate their mode of action. This would reveal the health benefits of telomerase activation through natural molecules and would shed new light onto the treatment of aging‑related diseases.
Introduction
Several studies have indicated that short telomere length is associated with aging-related diseases, including cardiovascular diseases (CADs), stroke, cancer, arthritis, osteoporosis, cataracts, diabetes type 2, hypertension, mental diseases, chronic obstructive pulmonary disease (COPD) and dementia (1). Telomere shortening can be affected by environmental factors, including physical activity, body mass index (BMI), hormone replacement therapy, smoking, chronic inflammation, oxidative stress, dietary antioxidants and vitamins (2–5). For instance, DNA-damage caused by various environmental factors triggers a DNA-damage response at telomeres that protects them from instability and shortening (6,7). Moreover, Vakonaki et al demonstrated an association between telomere length and drug abuse, which leads to premature biological aging (8). Telomere length has been proposed to be a biomarker of somatic cell aging, while the rate of increase of short telomeres has been linked to longevity in mammals (9). Indeed, when the length of the telomeres shortens below a threshold limit, cell growth is restricted and cells undergo cellular senescence or apoptosis (10). In a recent study, it was found that the administration of nutraceutical supplements may be linked to sustaining the telomere length in healthy adults (11). To determine the rate of telomere shortening and increase in the percentage of short telomeres with aging, we generated ‘BIOTEL version 2.4’ that was validated using data from Telomere Length Database Project (TLDP) (12), and allows the easy production of graphs and track telomere shortening in response to stimuli.
The shortening of telomeres can be reversed by the enzyme telomerase, which is active in high-proliferating cells, such as in male germ cells, activated lymphocytes, stem cells and cancer cells (13,14). It consists of two domains, namely a reverse transcriptase catalytic subunit (TERT) and an associated telomerase RNA component (TERC) (15). However, the majority of adult human somatic cells are telomerase-deficient and their proliferation contributes to progressive telomere shortening with age, ultimately leading to aging and death(16). In addition, telomerase-related gene mutations result in the development of certain diseases, such as Dyskeratosis Congenita (DKC) that is the first disease to be associated with mutations in human telomerase gene (17). Telomerase mutations have also been detected in aplastic anemia, Hoyeraal-Hreidarsson syndrome and idiopathic pulmonary fibrosis, while numerous epidemiological studies have demonstrated that telomerase activity is associated with pregnancy complications (18,19) and mental disorders (20). Thus, based on all the above, telomerase activators may be potent agents in anti-aging and in the treatment of telomerase-dependent diseases. It has been further demonstrated that telomerase activators enhance the efficiency of the DNA repair process and protect cells from stress and DNA-damaging conditions (21). Telomerase activation has been achieved through natural molecules, synthetic molecules and genetic manipulation and intervention (22). Several extracts from the Astragalus membranaceus root have been studied as possible telomerase activators (22–26). Such an extract is TA-65, a natural product telomerase activator marketed since 2008, that has been found to lengthen telomeres in humans (23). A previous in vitro study on human CD4 and CD8 T-cells suggested that cycloastragenol (CAG), a triterpenoid saponin compound obtained from Astragaloside IV hydrolysis that is the main compound in Astragalus, increased telomerase activity and reduced the effects of aging (24). Product B, a herb nutraceutical that contains ‘telomere support’ compounds and antioxidants, has also been suggested to be a potent telomerase activator, although no long-term test data are currently available.
The aim of the present study was to test supplements and natural extracts for their capacity to enhance telomerase activity in human peripheral blood mononuclear cells (PBMCs). We demonstrate that Centella asiatica extract formulation (08AGTLF) can lead to significantly higher telomerase activation compared to untreated cells, as well as TA-65 and other supplements containing Astragalus extract and CAG. This is the first study, at least to our knowledge, to demonstrate that a natural formulation, such as Centella asiatica extract formulation (08AGTLF) can lead to such high telomerase activity relative to control cells.
Materials and methods
Formulations
Centella asiatica extract formulation (08AGTLF) which consisted of >95% high-purity triterpenes was obtained from ApexBio Company. Oleanolic acid (OA) was obtained from Sigma-Aldrich and maslinic acid (MA) was obtained from Extrasynthese. Nutrient 1 and Nutrient 2 (contents shown below) were obtained from Lumis Research S.A. Nutrient 3 and Nutrient 4 (contents shown below) were obtained from Natural Doctor S.A. Each compound was dissolved in ethanol to achieve various concentrations to be tested in the cell cultures.
Cell isolation and telomerase activity measurements
The protocol of this study was approved by the Ethics Committee for Patients and Biological Material of the University of Crete with reference no. 63/22.03.2019. All procedures performed involving human participants were carried out under the ethical standards of the 1964 Helsinki declaration and its later amendments, or comparable ethical standards. The study was performed using samples prepared from healthy donors that volunteered to participate in the study. The samples were anonymized and personal data were managed according to the EU General Data Protection Regulation (GDPR).
PBMCs where isolated from the blood samples by Ficoll-Hypaque gradient centrifugation. The cells were grown in DMEM (Biochrom AG; F0455) supplemented with 10% fetal bovine serum (FBS; 10500-064, heat-inactivated; Invitrogen; Thermo Fisher Scientific), glutamine (4 mM; Biosera XCT1715), gentamycin (15710–049; Invitrogen; Thermo Fisher Scientific) and penicillin/streptomycin (100 U/ml; Biosera LMA4118). Prior to the addition of the test agents, the cells were cultured in serum-free medium for 24 h at 37°C and 5% CO2. The PBMCs were then treated with the compounds at various concentrations, for 24–72 h. PBMCs samples were collected at 24–72 h following treatment, washed in PBS buffer and stored at −80°C. Telomerase activity was measured using a commercial telomerase PCR-ELISA (Sigma-Aldrich), based on the telomeric repeat amplification protocol, as previously described (27–30). All treatments for each condition were performed in triplicates.
Contents of Nutrients 1, 2, 3 and 4
Nutrient 1 (My Shape) contained the following: Alpha lipoic acid, cinnamon) bark dry extract 1/4 (Cinnamomum zeylanycum N.), magnesium citrate, L-glutamine, L-carnitine tartrate, potassium citrate, ascorbic acid, magnesium ascorbate, green tea (Camellia sinensis K.) leaves dry extract titrated to 95% polyphenols, natural vitamin E acetate 50%, enzimix (amylase, protease, glucose amilase, lipase, cellulase, lactase, pectinase), niacin, bitamin B1, vitamin K2 Mena Q7 0.2%, selenium methionine, vitamin B2, β-carotene, vitamin B5, choline bitartrate, inositol, para-aminobenzoic acid (PABA), vitamin B6, vitamin B12 1%, chromium picolinate, vitamin D3 2.5%, biotin, folic acid, anti-caking agent (cellulose, mono- and diglycerides of fatty acid, magnesium stearate, silica dioxide).
Nutrient 2 (My Health) contained the following: Mix Vitamin (ascorbic acid, vitamin E acetate 50% natural, niacin, vitamin B1, vitamin K2 Mena Q7 0.2%, vitamin B6, β-carotene, vitamin B12 1%), anti-caking agents (microcrystalline cellulose, mono- and diglycerides of fatty acids, magnesium stearate, silica dioxide).
Nutrient 3 (Vit. D3&K2 Cofactors, 1 capsule) contained the following: 2,000 OH25D3, 100 µg vitamin K2 (MK7), 56 mg elemental magnesium as magnesium bisglycinate.
Nutrient 4 (REYOUTH, 1 capsule) contained the following: Vitamin C (50 mg), Magnesium (58 mg), CAG (16 mg) and amino-acids mix containing L-glutamine, L-lysine, L-proline, L-glycine, L-arginine, L-leucine, L-histidine, L-isoleucine, L-valine, L-methionine, L-tyrosine, glutamic acid, L-phenylalanine, L-serine, L-threonine, L-alanine, L-citrulline, L-taurine, L-tryptophan, aspartic acid, Vitamin E (12 mg), calcium, Vitamin B3, Broccoli dry extract, dry fruit and vegetable extract, blend of digestive enzymes (Enzymix), potassium, Vitamin B1, Zinc, Vitamin B6, manganese, phosphorus, B-carotene, Vitamin B5, inositol, Vitamin K2, Vitamin B2, PABA, Vitamin D3, biotin, chromium, copper, selenium, molybdenum, Vitamin B12.
TA-65 (1 capsule) contained the following: Astragalus membranaceus moench extract (TA-65®MD, 8 mg).
Statistical analysis
The mean (xm), standard deviation (SD) and the estimated approximated 95% confidence interval (95% CI) (xm±1.96 SD/√n (where n -n=3- was the number of replications) of the absorbance values were applied. All experiments were performed in triplicates and the mean values were used for the data presentation of differences of telomerase activity triggered by each formulation vs. the control, expressed as P-values resulting from one-way ANOVA followed by Dunnett's post-hoc test for pairwise comparisons with untreated cells. All statistical analyses were performed in IBM SPSS Statistics 24.0 and diagrams were created using Excel 365 for Windows (Microsoft Corp.) and a value of P<0.05 was considered to indicate a statistically significant difference.
Results
A summary of the compounds used and the calculated concentrations (µg/ml) in vitro is presented in Table I. Fig. 1 depicts the mean values of the telomerase activity (expressed in absorbance units, A450nm-A690nm) of cells treated with the formulations and compounds (08AGTLF, TA-65, Nutrient 4, OA and MA) in comparison with the ethanol-only treated cells, hereafter referred as untreated cells. Importantly, all the compounds tested were not toxic for the cells, as they only led to small amount of apoptosis or necrosis (13–15%), similar with the untreated cells (13%; data not shown). 08AGTLF exhibited the highest telomerase activity, 1.35 absorbance units (95% CI, 1.154–1.546) at the concentration of 0.02 µg/ml and 1.18 (95% CI, 1.088–1.278) at the concentration of 0.2 µg/ml, while it decreased at the concentration of 2 µg/ml. The difference in telomerase activity reached the levels of 8.8-fold increase relative to the untreated cells at the concentration of 0.02 µg/ml. Importantly, the differences in telomerase activity of the treated cells compared to the untreated ones were statistically significant with P-values <0.001 and <0.0001 for the 0.02 and 0.2 µg/ml concentrations, respectively.
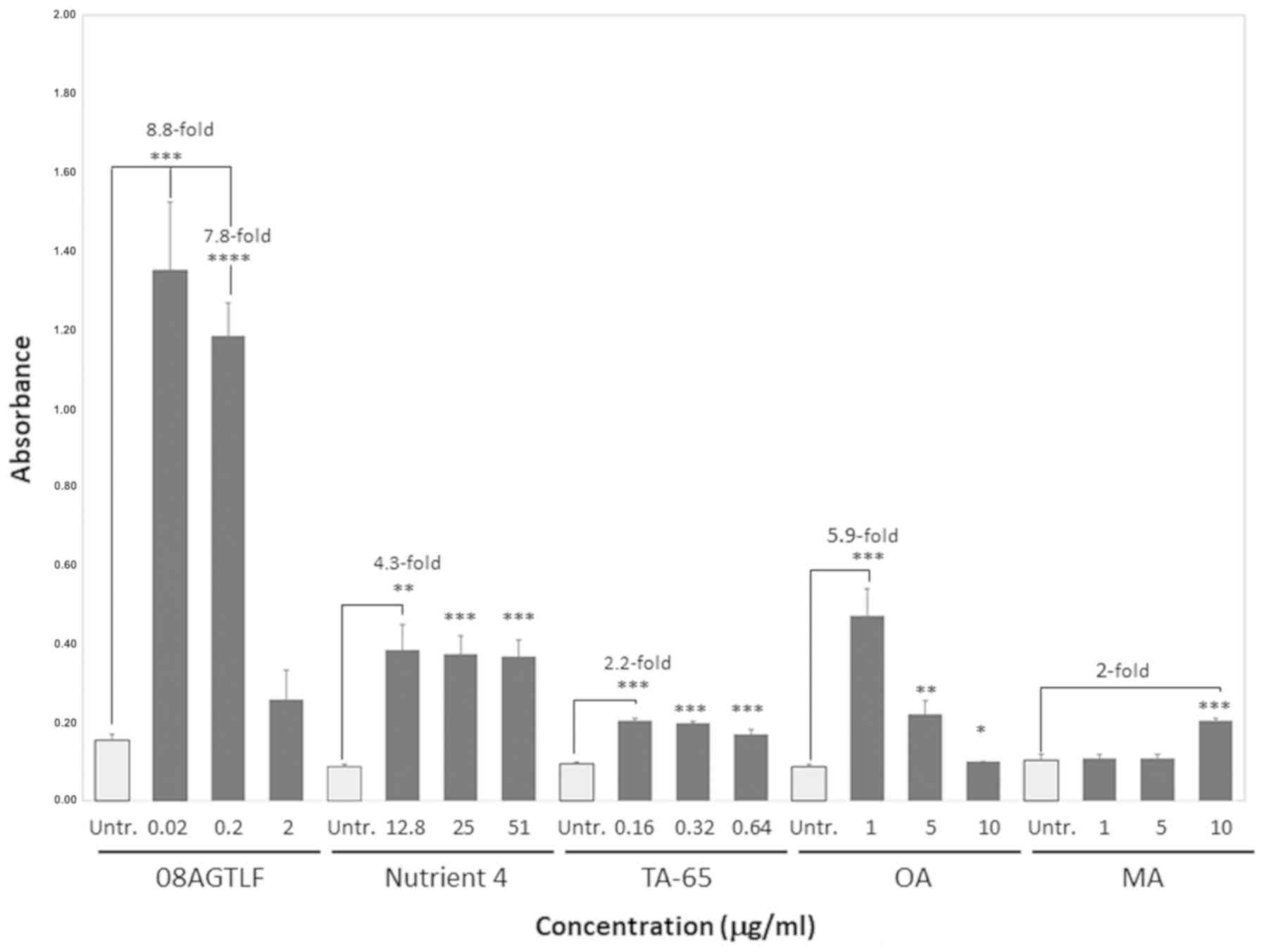
Figure 1. - Telomerase activity expressed in absorbance units (A450nm-A690nm) in the untreated cells or after treatment of PBMCs with 08AGTLF (Centella asiatica extract folmulation), Nutrient 4, TA-65, OA (oleanolic acid) and MA (maslinic acid). Error bars represent the standard deviation of the mean. Each mean was estimated from triplicate experiments. Asterisks indicate significant differences in the mean absorbance values measured after treatment with each activator compared with untreated cells (*P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001) at the indicated concentrations.
.../...
.