.
S O U R C E : International Journal of Molecular Sciences
Abstract:
Aging is a natural phenomenon that occurs in all living organisms. In humans, aging is associated with lowered overall functioning and increased mortality out of the risk for various age-related diseases. Hence, researchers are pushed to find effective natural interventions that can promote healthy aging and extend lifespan. Royal jelly (RJ) is a natural product that is fed to bee queens throughout their entire life. Thanks to RJ, bee queens enjoy an excellent reproductive function and lengthened lifespan compared with bee workers, despite the fact that they have the same genome. This review aimed to investigate the effect of RJ and/or its components on lifespan/healthspan in various species by evaluating the most relevant studies. Moreover, we briefly discussed the positive effects of RJ on health maintenance and age-related disorders in humans. Whenever possible, we explored the metabolic, molecular, and cellular mechanisms through which RJ can modulate age-related mechanisms to extend lifespan. RJ and its ingredients—proteins and their derivatives e.g., royalactin; lipids e.g., 10-hydroxydecenoic acid; and vitamins e.g., pantothenic acid—improved healthspan and extended lifespan in worker honeybees Apis mellifera, Drosophila Melanogaster flies, Gryllus bimaculatus crickets, silkworms, Caenorhabditis elegans nematodes, and mice. The longevity effect was attained via various mechanisms: downregulation of insulin-like growth factors and targeting of rapamycin, upregulation of the epidermal growth factor signaling, dietary restriction, and enhancement of antioxidative capacity. RJ and its protein and lipid ingredients have the potential to extend lifespan in various creatures and prevent senescence of human tissues in cell cultures. These findings pave the way to inventing specific RJ anti-aging drugs. However, much work is needed to understand the effect of RJ interactions with microbiome, diet, activity level, gender, and other genetic variation factors that affect healthspan and longevity.
1. Introduction
Reduced mortality from both communicable and noncommunicable diseases accelerates the growth of aging populations worldwide. However, long living entails reduced healthspan out of increased burden of noncommunicable diseases [1]. Aging, healthspan, and lifespan are tightly related concepts. Though aging is a natural phenomenon, numerous factors such as stress, poor nutrition, and pollution are associated with increased internal production of free radicals, which enhance chronic subclinical inflammation and lead to faster aging [2,3]. Aging-related oxidative damage and inflammation contribute to a trail of cellular and molecular alterations: telomere attrition, epigenetic alterations, genome instability, reduced proteostasis, disturbed nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication. In this respect, aging represents a process of random cellular degradation [4,5,6], which is associated with increased likelihood of progressive loss of function, decreased fertility, and increased risk of various diseases (both physical and cognitive)—contributing to premature death and increased morbidity in aging populations (Figure 1) [2,7].
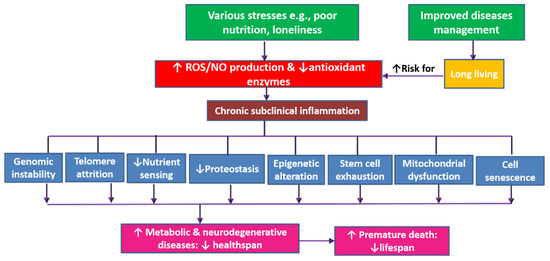
Figure 1. The dynamics of aging-related decrease of healthspan and lifespan. The production of free radicals increases with aging (which is on the rise out of improved disease management) and also with exposure to various stresses (e.g., pollution, poor nutrition, and psychological stress) leading to oxidative stress. This is associated with reduced production of antioxidant enzymes and activation of inflammatory pathways. Both oxidative stress and chronic inflammation lead to a trail of cellular and molecular alterations: telomere attrition, epigenetic alterations, genome instability, reduced proteostasis, disturbed nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication. Such age-related physiological alterations result in poor health and loss of function out of increased occurrence of various metabolic and neurodegenerative disorders. Age-related disorders are associated with increased mortality and premature death i.e., decreased lifespan.
A recent review indicates that family profiles of centenarians emphasize the contribution of “protective or longevity genes” to extreme longevity as well as to low incidence of age-related diseases in this group [5]. Apart from genetic factors, favorable environmental conditions and healthy lifestyles contribute to low morbidity in people who survive to extreme ages [8,9]. Therefore, the worldwide substantial increase of aging populations, who encounter a trail of debilitating illnesses, has driven researchers to find strategies to slow aging and support a healthy lifespan by delaying the onset of age-related diseases [5]. Research denotes that the natural decline of function that occurs with aging, as well as age-related disorders, can be prevented and/or reversed through environmental modifications such as dietary interventions [10,11]. Accumulating evidence indicates that functional foods can promote healthy aging, reduce morbidity, and lengthen healthy lifespans [12]. Royal jelly (RJ) is considered a functional food (with a documented safety profile) because it has a range of pharmacological activities: antioxidant, anti-inflammatory, antitumor, antimicrobial, anti-hypercholesterolemic, vasodilative, and hypotensive. Indeed, RJ has been widely used to treat several health conditions such as diabetes mellitus, cardiovascular diseases, and cancer, to name a few [13,14].
RJ is a creamy substance secreted from the mandibular and hypopharyngeal glands of the worker honeybee Apis mellifera. It is the food of all bee larvae during their first 3 days of life; workers then shift to worker jelly (WJ), composed mainly of honey and pollen, while bee queens continue to consume RJ [15,16,17]. It is suggested that RJ is a potent promoter of healthy aging and longevity because it enhances overall health and fertility of queen bees, who may lay up to 3000 eggs a day and survive as long as five years compared with infertile workers that live up to 45 days only [18,19,20]. However, studies of the effect RJ on longevity in humans are scarce, presumably due to technical and ethical reasons. In this respect, this paper reviews, synthesizes, and discusses the most relevant studies that used RJ to enhance healthspan and extend lifespan in different species. We also examined the metabolic, molecular, and cellular mechanisms associated with the longevity-promoting properties of RJ.
2. Chemical Composition of RJ
Water constitutes 60–70% (w/w) of fresh RJ; pH of fresh RJ usually ranges between 3.6 and 4.2. The various pharmacological properties of RJ are attributed to its unique and rich composition of proteins, carbohydrates, vitamins, lipids, minerals, flavonoids, polyphenols, as well as several biologically active substances [21]. Herein, we provide a detailed description of most ingredients of RJ.
2.1. Sugars
Sugars comprise 7.5–15% of RJ content. Fructose and glucose constitute 90% of the total sugar fraction of RJ, whereas sucrose accounts for 0.8–3.6%. RJ contains very small amounts of other sugars such as maltose, trehalose, melibiose, ribose, and erlose [22]. RJ sugar content varies remarkably from one sample to another based on season, geographical location, botanical origin, bee species, and method of production. For example, French RJ contents of sucrose and erlose are less than 1.8% and 0.4%, respectively, whereas their concentrations in RJ produced by sugarcane feeding are comparatively higher (7.7% and 1.7%) [23]. Sugars of RJ are thought to contribute to its epigenetic effects, given that RJ sugar content is extremely high compared with WJ (the main food of bee workers). Meanwhile, supplementing WJ with fructose and glucose (4%) fosters the development of larvae to adult workers. In the same way, a gradual increase of WJ sugar concentrations (up to 20%) increases larval consumption of WJ and eventually results in the development of intercastes (midway between queens and workers) and queens at a rate similar to that obtained by pure RJ for in vitro rearing [24,25]. Thus, sugars of RJ represent a phagostimulant that functions through the insulin/insulin-like signaling cascades and the nutrient sensing mTOR pathway to derive larval development by increasing quantities of ingested food and increasing intake of nutrients necessary for queen development [25].
2.2. Lipids
Among the principal bioactive constituents of RJ, lipids constitute 7–18% of RJ content; 90% of these lipids are rare and unique short hydroxy fatty acids with 8–12 carbon atoms in the chain and dicarboxylic acids. The most prominent RJ fatty acids in order are 10-hydroxydecanoic acid (10-HDA), 10-hydroxy-2-decenoic acid (10H2DA), and sebacic acid (SA) [18]. The 10-HDA exerts epigenetic control over caste differentiation of Apis mellifera by inhibiting histone deacetylases, which catalyze the hydrolysis of ε-acetyl-lysine residues of histones [25,26]. Due to the low pH of RJ, 10-HDA acts as a strong bactericidal. It therefore, protects bee larvae against virulent bacterial infections that affect bee hives such as those caused by some strains of Paenibacillus larvae [27]. In mammals, it protects mice against pulmonary damage induced by lipoteichoic acid, a toxin from Staphylococcus aureus [28]. It also exerts an antibacterial effect against various pathogenic bacterial species in human cancer colon cells [29]. The 10-HDA may be used to treat age-related neurodegenerative disorders given its documented neurogenic activity—it stimulates neuronal differentiation from progenitor cells (PC12) cells through mimicking the effect of brain-derived neurotrophic factor [30]. In addition, it possesses neuroprotective effects against glutamate- and hypoxia-induced neurotoxicity [31]. The 10-HDA may also be used for manufacturing cosmetics and anticancer drugs, given that it increases skin-whitening and exerts antiproliferative effects on B16F10 melanoma cells by inhibiting the expression of microphthalmia-associated transcription factor and tyrosinase-related protein 1 (TRP-1) and TRP-2 [32]. Moreover, 10-HDA has been identified as an inhibiting factor of matrix metalloproteinases (MMPs)—proenzymes activated by proteolytic cleavage under inflammatory conditions, which degrade matrix and non-matrix proteins and contribute to tissue aging (e.g., skin) and cause serious disabling diseases such as rheumatoid arthritis [33,34].
SA, 10-HDA, and 10H2DA, demonstrate anti-inflammatory effects through regulation of several proteins involved in the mitogen-activated protein kinase (MAPK) and nuclear factor kappa-B signaling [29,35]. Moreover, these acids mediate estrogen signaling by enhancing the activity of estrogen receptors (ERs) ERα, ERβ [36], which can benefit bone, muscle, and adipose tissue in a sex-dependent manner [31]. A derivative of 4-hydroperoxy-2-decenoic acid known as 4-hydroperoxy-2-decenoic acid ethyl ester (HPO-DAEE) prevents 6-hydroxydopamine-induced cell death in human neuroblastoma SH-SY5Y cells through triggering slight emission of reactive oxygen species (ROS), which stimulates the production of antioxidants via activation of antioxidant pathways: nuclear factor erythroid 2 (NRF2)-antioxidant response element (ARE) and eukaryotic initiation factor 2 (eIF2α), an upstream effector of the activating transcription factor-4 (ATF4) [37]. HPO-DAEE also demonstrates anticancer effects through accumulation of intracellular ROS and activation of proapoptotic CCAAT-enhancer-binding protein homologous protein expression [38].
2.3. Proteins
Proteins are the dominant ingredient of RJ (50% of its dry matter) and more than 80% of total RJ proteins are composed of nine major RJ proteins (MRJPs, 49–87 kDa)—the first five MRJPs constitute up to 82–90% of MRJPs. Glycosylation and phosphorylation of MRJPs is essential for biological processes that involve glycoproteins, such as cell adhesion, cell differentiation, cell growth, and immunity. MRJPs modulate the development of female larvae, not only through their high nutritional value but mainly through physiological activity of their highly homologous 400–578 amino acids that contribute to RJ’s role in cell proliferation, cytokine suppression, and antimicrobial activity [12].
Research documents anti-senescence activity of MRJPs for human cells in vitro [39]. MRJP1 is the most dominant among all MRJPs; essential amino acids constitute 48% of its content. Circular dichroism measurements indicate that the secondary structure of MRJP1 consists of 9.6% α-helices, 38.3% β-sheets, and 20% β-turns [22]. MRJP1 exists in two distinct forms: oligomer and monomer. Oligomer MRJP1 is highly heat-resistant and it is considered a predominant proliferation factor compared with MRJP2 and MRJP3 [40]. High-performance liquid chromatography and SDS-PAGE analyses of MRJP1 revealed the presence of a 57 kDa monomeric glycoprotein, which can be degraded at 40 °C, known as royalactin. Royalactin mimics the effect of epidermal growth factor (EGF) in rat hepatocytes and modulates the development of bee larvae [41]. Royalactin is reported to bind with the most sensitive regions in mouse embryonic stem cell culture, resulting in activation of a pluripotency gene network that enables self-renewal of stem cells [42]. MRJP1 exerts nematicidal activity against C. elegans via constant downregulation of a rate-controlling enzyme of the citric acid cycle known as isocitrate dehydrogenase encoding the idhg-1 gene [43]. On the other side, MRJP2 and its isoform X1 exhibit potent anticancer effects and protect hepatocytes against CCl4 toxicity by inducing caspase-dependent apoptosis, scavenging intracellular free radicals, inhibiting tumor necrosis factor (TNF)-α, and mixed lineage kinase domain-like protein [44].
RJ contains proteins other than MRJPs, albeit in small amounts, such as royalisin, jelleines, and aspimin. Royalisin and jelleines are common RJ antimicrobial peptides that enhance efficiency of the immune response of bee larvae to various infections. The structure of royalisin is highly compact due to its high cysteine content, which boosts its stability at low pH and high temperature. On the other hand jelleines are thought to stem from trypsin digestion of MRJP1 by the action of exo-proteinase of the hypopharyngeal glands on C-terminal to N-terminal tryptic fragment. Peptides of royalisin and jelleines possess hydrophobic residues, which contribute to their antimicrobial properties by affecting functions of bacterial membranes. RJ also contains apolipophorin III-like protein, a lipid binding protein that exerts antimicrobial effect by carrying lipids into aqueous environments through the formation of protein–lipid complexes. In addition, the antibacterial effect of RJ is partially attributed to its glucose oxidase enzyme, which catalyzes the oxidation of glucose to hydrogen peroxide [45]. Recently, examination of whole RJ and single protein bands by off-line LC-MALDI-TOF MS glycomic analyses, complemented by permethylation, Western blotting, and arraying data, revealed the presence of glucuronic acid termini, sulfation of mannose residues, core β-mannosylation of the N-glycans, and a fairly scarce zwitterionic modification with phosphoethanolamine, which may contribute to the development of honey bees and their innate immunity [46].
2.4. Phenols, Flavonoids, and Free Amino Acids
The antioxidant potency of RJ, at least in part, is attributed to its polyphenolic compounds and flavonoids, which are measured based on gallic acid and rutin equivalent (GAE and RE), respectively [47]. RJ contains 23.3 ± 0.92 GAE µg/mg and 1.28 ± 0.09 RE µg/mg of total phenolics and flavonoids, respectively. The vast phenolic content of RJ consists of pinobanksin, organic acids (e.g., octanoic acids, dodecanoic acid, 1,2-benzenedicarboxylic acid), and their esters. Meanwhile, flavonoids of RJ can be differentiated into four groups: (1) flavanones e.g., hesperetin, isosakuranetin, and naringenin; (2) flavones e.g., acacetin, apigenin and its glucoside, chrysin, and luteolin glucoside; (3) flavonols e.g., isorhamnetin and kaempferol glucosides; and (4) isoflavonoids e.g., coumestrol, formononetin, and genistein. The antioxidant activity of these components contributes to the antiapoptotic and anti-inflammatory properties of RJ [18]. Age of larvae that produce RJ (1–14 days) and harvesting time affect RJ content of phenols and amino acids; RJ harvested from the youngest larvae (one-day-old) within 24 h contains higher proteins and polyphenolic compounds and exhibits stronger free radical scavenging effect compared with RJ harvested from older larvae or later than 24 h [47]. Small peptides such as di-peptides (Lys-Tyr, Arg-Tyr, and Tyr-Tyr) obtained from RJ proteins hydrolyzed by protease N possess high antioxidative activity owing to their phenolic hydroxyl groups, which scavenge free radicals by releasing a hydrogen atom [48].
RJ is rich in amino acids, including essential ones [22]. Concentrations of free amino acids in RJ increase with harvesting time from 4.30 mg/g at 24 h to 9.48 mg/g at 72 h, whereas levels of total amino acids decrease by time from 197.96 mg/g at 24 h to 121.32 mg/g at 72 h [49]. LC/MS method analysis and hydrophilic interaction liquid chromatography-tandem mass spectrometry indicate that lysine is the most prominent free amino acid in RJ (62.43 mg/100 g), followed by proline (58.76 mg/100 g), cystine (21.76 mg/100 g), and aspartic acid (17.33 mg/100 g). RJ contains less than 5 mg/100 g of other amino acids such as valine, glutamic acid, serine, glycine, cysteine, threonine, alanine, tyrosine, phenylalanine, hydroxyproline, leucine-isoleucine, and glutamine [18,50].
The amino acid content in protease-treated RJ (pRJ)—obtained by removal of two allergen proteins to convert RJ hydrolysates into shorter chain monomers that are easy to absorb—is greater than in crude RJ [51]. Moreover, eluted water pRJ with 30% MeOH contains higher levels of dipeptides and tripeptides than pRJ. These components are likely to possess a lifespan-prolonging activity since pRJ increases lifespan of C. elegans compared with RJ [52]. Evidence signifies that amino acids of RJ can prolong lifespan in mammals. Long-term dietary supplementation of branched-chain amino acid-enriched mixture (BCAAem), which contains 3 branched-chain amino acids that can be found in RJ (leucine, isoleucine, and valine), enhances mitochondrial biogenesis and sirtuin 1 expression, and reduces ROS production in cardiac and skeletal muscle, which is associated with alleviation of age-related muscle dysfunction, resulting in an increase of the average lifespan of male mice. The effect of these amino acids on mitochondrial biogenesis involved activation of signaling pathways of endothelial nitric oxide synthase (eNOs) and mTOR and its substrates: S6K and eukaryotic translation initiation factor (eIF4E)-binding protein (4E-BP1) [53].
2.5. Vitamins, Minerals, and Bioactive Substances
Pantothenic acid (vitamin B5) is the most abundant vitamin in RJ (52.8 mg/100 g), followed by niacin (42.42 mg/100 g). RJ contains small amounts of various B group vitamins (B1, B2, B6, B8, B9, and B12), ascorbic acid (vitamin C), vitamin E, and vitamin A [18,22]. Gardner (1948) noted that pantothenic acid of RJ is a lifespan-extending agent [54].
Mineral salts constitute 1.5% of RJ content [18]. Inductively coupled plasma optical emission spectroscopy and double focusing magnetic sector field inductively coupled plasma mass spectrometry indicate that RJ contains small amounts of various minerals and trace elements such as K, Na, Mg, Ca, P, S, Cu, Fe, Zn, Al, Ba, Sr, Bi, Cd, Hg, Pb, Sn, Te, Tl, W, Sb, Cr, Mn, Ni, Ti, V, Co, and Mo. Whereas concentrations of trace and mineral elements in honey vary according to botanical origin, RJ content of trace elements and minerals is highly constant. In this respect, RJ can be considered a form of larval lactation that possesses homeostatic adjustment, same as mammalian and human breast milk [55].
RJ contains high amounts of acetylcholine (Ach, 4–8 mM), and RJ concentration of Ach is highly conserved because of its acidic pH [56]. It is well-known that Ach acts as a neurotransmitter that plays a major role in memory formation and cognitive functioning. Glucose metabolism and insulin contribute to Ach synthesis by controlling the activity of choline acetyltransferase [57]. In this respect, consumption of RJ may prevent the development of cognitive dysfunction thanks to its Ach content. In addition, Ach content of RJ has a survival promoting effect [56].
RJ is rich in nucleotides such as free bases (e.g., adenosine, uridine, guanosine, iridin, and cytidine) and phosphates (e.g., adenosine diphosphate (ADP), adenosine triphosphate (ATP), and adenosine monophosphate (AMP)). Nucleotides constitute 2682.9 mg/kg and 3152.8 mg/kg in fresh and commercial RJ, respectively. Levels of ADP, ATP, and AMP are higher in fresh RJ, and therefore, they can signify RJ freshness. These compounds are necessary for organisms’ physiological activities e.g., metabolic degradation of intracellular ATP is essential to provide cells with energy necessary for transport systems and enzymatic activities of proteins [22,58]. Among all nucleotides, AMP N1-oxide is considered a unique active component that exists nowhere in nature except in RJ. AMP N1-oxide demonstrates neurogenic and neurotrophic activities: it stimulates neurite outgrowth and induces differentiation of PC12 cells into neurons similar to sympathetic neurons. This action is similar to that of nerve growth factor, which functions through activation of two cascades of cellular signaling MAPK/extracellular signal-regulated kinase 1 or 2 (ERK1/2) and phosphatidylinositol 3-kinase/Akt pathways. The neurite outgrowth-promoting activity of AMP N1-oxide is mediated by adenylate cyclase-coupled adenosine A2A receptors, which are highly expressed in the brain (striatum in particular)—adenosine A2A receptors prevent radical formation and apoptosis, and they contribute to early neuronal development and regulation of synaptic plasticity [18,59].
3. Healthspan and Longevity Effects of RJ in Various Species
3.1. RJ Enhances Fertility and Longevity in Population of the Beehive
Numerous bioactive elements are abundant in RJ, making it an optimal food and a well-balanced nutrient-rich diet [25] that, when consumed by bee larvae, induces their development into queens. Meanwhile, larvae that consume honey or pollen grow into workers, which live shorter and are unable to reproduce. On the other hand, emerging bee workers fed RJ-rich diet acquire queen-like morphogenic characteristics such as increased body size and ovary development [41,60,61]. Real-time RT-PCR and HPLC-ECD analysis denoted that augmentation of fertility (ovarian development) in bee workers by RJ is the result of enhancement of brain levels of tyrosine, dopamine, and tyramine by RJ [61], as well as activation of epidermal growth factor receptor (EGFR), which increases the production of the juvenile hormone, titre, known to regulate growth [41]. RJ also improves the memory of bee workers [62] and increases their survival [15,41,56,62,63], owing to its high Ach concentration [56], MRJP1 content of royalactin [41], MRJPs 2, 3, and 5 [25], and a water soluble RJ protein extracted by precipitation with 60% ammonium sulfate (RJP60) [15].
3.2. RJ Enhances Healthspan and Longevity in Other Species
Based on the assumption that RJ is the main factor contributing to long survival of bee queens, several studies examined the lifespan-expanding effects of RJ in diverse species (Figure 2), and results seem to be consistent with the naturally occurring model of bee queens.
Figure 2. Royal jelly and its components improve healthspan and extend lifespan in different species.
3.2.1. Drosophila Melanogaster (Fruit Fly)
The fruit fly Drosophila melanogaster (M.) has been widely used in the literature as an invertebrate model to understand the pathology of numerous diseases and to examine the effect of various agents, including RJ. The earliest study of the longevity effect of RJ dates back to 1948. That study reported that large doses of dehydrated RJ, its pantothenic acid, as well as its water soluble and insoluble organic acids significantly extended lifespan of Drosophila M. compared with controls. The author suggested that pantothenic acid of RJ may have an anti-aging effect by itself or by synergizing the action of other vitamins [54]. In a relatively recent study, RJ (0.1, 0.3, and 0.5 g) significantly increased the average lifespan of both male and female flies—the effect was attributable to enhancement of the antioxidation capacity noticed in flies treated by high doses of RJ: increased superoxide dismutase (SOD) and catalase levels [64].
As a replication of his experiment of RJ and royalactin in bees, Kamakura (2011) treated Drosophila Canton-S. larvae with royalactin and 20% fresh RJ. RJ treatment increased body size, cell size, and fertility; prolonged lifespan; and shortened developmental time from larva to adult compared with control. Same as in bees, RT-PCR and enzyme immunoassay showed that both RJ and royalactin induced EGFR signaling (not insulin signaling), which activated S6K in the fat body, which further increased the body size. EGFR also activated the MAPK pathway, which increased the synthesis of a biologically active ecdysteroid known as 20-hydroxyecdysone (20E) and juvenile hormone—an effect that was demonstrated by growth regulation—reducing the developmental time [41]. A subsequent study stated that supplementing Drosophila M. with 1% freeze-dried RJ (FDRJ) powder shortened developmental time, prolonged the lifespan of adult males, and increased females’ egg production without any morphological changes. In female flies, RT-PCR indicated that FDRJ was significantly associated with heightened gene expression of an insulin-like peptide known as dilp5, its insulin receptor (InR), and the nutrient sensing molecule mTOR, the mechanistic target of rapamycin—all these molecules are known to affect growth and reproduction. However, stimulation of the insulin/TOR signaling pathways was not associated with extension of lifespan of FDRJ-fed female flies [17]. Another study noted that supplementation of MRJPs, especially MRJP 1 and MRJP3, at an optimal dose of 2.5% (w/w) of diet significantly lengthened the mean lifespan of both male and female Drosophila. The longevity effect of MRJPs was positively associated with increased feeding and fertility. Microarray data and gene ontology enrichment analyses revealed that the molecular mechanism underlying increased lifespan and fertility was similar to that discovered by Kamakura (2011): MRJP supplementation upregulated the gene expression of S6K, MAPK, and EGFR in EGFR-mediated signaling. In addition, MRJPs improved the anti-oxidation capacity of flies by increasing the expression of CuZn-SOD gene i.e., SOD levels were higher, while malonaldehyde (MDA) levels were lower than control flies [12]. Another study replicated the size/growth enhancing effect of RJ at low concentrations (10–30%) in Drosophila M. [16]. However, males’ lifespan decreased by 20% RJ treatment whereas none of these concentrations affected the lifespan of females. Findings provided no evidence of RJ activation of insulin and EGFR signaling pathways, yet RJ regulated the gene expression related to oxidative stress and catabolism. The authors attributed the discrepancy noted between their results and findings of other studies to employing a slightly different strain of Drosophila (Canton-S), using a commercially available source of RJ, and difference in the nutrient contents of the control culture medium. On the other hand, higher concentrations (40–70%) of RJ had adverse effects: prolonged development time, shortened lifespan, increased mortality, and reduced productivity in both sexes. Data on global gene expression indicated that excess nutrients in high doses of RJ altered cellular processes as a result of altering genes involved in amino acid metabolism and encoding glutathione S transferases, which detoxify xenobiotic compounds [16].
3.2.2. Gryllus Bimaculatus Cricket and Silkworms
The two-spotted cricket Gryllus (G.) bimaculatus, a member of primitive group Polyneoptera, is another kind of species that were used to examine the longevity effect of RJ. RJ dietary supplementation to G. bimaculatus during early nymph stage significantly decreased developmental time, extended lifespan, and increased the body size of both males and females in a dose-dependent manner (8−15% w/w) compared with the control high protein, sugar, and lipid diet. The prolonged lifespan of RJ was not due to an extended nymph stage since the adult stage emerged earlier in crickets fed RJ than those fed the control diet. Similarly, RJ administration increased body size and egg size in female silkworms. The authors concluded that the effects of RJ were not attributed to the nutritional supplement itself; however, they did not examine the molecular mechanism behind them [65].
3.2.3. Caenorhabditis (C.) Elegans Nematodes
Caenorhabditis (C.) elegans nematodes have been used in several instances to test the longevity-promoting activity of RJ. Japanese researchers found that RJ, protease-treated RJ (pRJ), pRJ-Fraction 5 (pRJ-Fr.5), and a derivative of pRJ-Fr.5—10-HDA (the main lipid of RJ)—extended the lifespan of C. elegans. RJ 10 µg/mL was an optimal dose for enhancing longevity by 7−9%, while RJ 1 or 100 µg/mL had no effect. Meanwhile, all pRJ concentrations significantly prolonged the mean lifespan—though the greatest effect was noticed at 10 µg/mL, which increased mean lifespan by 7–18%. The longevity effect of combined pRJ-Fr.5 and 10-HDA was greater than that induced by each treatment on its own. DNA microarray and RT-PCR showed that the longevity-promoting effect was attributed to reduction of the insulin/IGF-1 signaling—pRJ-Fr.5 upregulated the expression of dod-3 gene and downregulated the expression of ins-9, an insulin-like peptide gene, along with dod-19, dao-4, and fkb-4 genes (further details are shown below in the mechanism section) [52]. In two subsequent studies RJ, pRJ, and 10-HDA enhanced longevity and increased stress resistance of C. elegans against thermal, irradiation, and oxidative stress [66,67]. Intact, deglycosylated, and mildly heat-treated royalactin extended mean lifespan of C. elegans by 18–34%—higher concentrations produced the vastest lifespan-extending effect. Royalactin also enhanced locomotion, which indicates promotion of healthy aging [68].
3.2.4. Mice
Few studies investigated the lifespan extending activity of RJ in mice. In an early study, intermediate and high doses (50 and 500 ppm) of RJ significantly prolonged the mean lifespan of C3H/HeJ mice by 25%, whereas RJ at a low dose (5 ppm) yielded no significant effect. Meanwhile, all doses of powdered RJ, contrary to bee and Drosophila studies, had no effect on mice growth, food intake, or appearance compared with control mice. RJ treatment significantly lowered kidney DNA and serum levels of 8-hydroxy-2-deoxyguanosine, a marker of oxidative stress that increases with aging [2]. Similarly, long-term intragastric administration of RJ and pRJ to a d-galactose-induced aging mice model resulted in numerous anti-aging and healthspan effects: preventing aging-related weight loss, improving memory and motor performance, and delaying aging-related atrophy of thymus, thus preventing diminution of the immune function compared with control animals. The effects were attributed to inhibition of lipid peroxidation and improvement of levels of antioxidant enzymes [69]. Likewise, dietary supplementation of RJ and pRJ (0.05% or 0.5%, v/v) to genetically heterogeneous head tilt mice—which exhibit vestibular dysfunction, imbalanced position, and inability to swim—could not prolong lifespan but significantly delayed age-related impairment of motor functions, positively improved physical performance of treated mice on four tests (grip strength, wire hang, horizontal bar, and rotarod), lowered age-related muscular atrophy, increased markers of satellite cells (muscle stem cells), and suppressed catabolic genes [70]. Another study examined the survival-expanding time (not lifespan extension) of oral RJ treatment (75, 150, and 300 mg/kg body wt/day for 13 consecutive days) following NaNO2 intraperitoneal injection or decapitation as models of brain hypoxia and complete brain ischemia [71]. Findings indicated that the intermediate dose of RJ (150 mg) significantly expanded survival time, whereas RJ 75 mg had no significant effect, meanwhile RJ 300 mg significantly decreased survival time. The author suggested that low pH of RJ (3.6 to 4.2) in mice treated with high doses of RJ induced activation of acid-sensing channels, increased acidosis of extracellular fluid, and aggravated brain ischemia [71].
4. RJ Might Enhance Longevity in Humans by Promoting General Health
It is an important question whether observed effects of RJ on healthspan and longevity in model organisms (e.g., bees, fruit flies, C. elegans, silkworms, crickets, and mice) can be generalized to humans. A small number of in vitro studies examined the antiaging activity of RJ, 10-HDA, and MRJPs on human cell lines, and results support findings reported from studies of model organisms. In two experiments, normal human skin fibroblasts were ultraviolet-irradiated (as a model of skin photoaging) and treated with RJ and 10-HDA. Results revealed that RJ and 10-HDA protected cells against ultraviolet A- and B-induced ROS-related oxidative damage, decreased cellular senescence, stimulated the production of procollagen type I and transforming growth factor-β1 [33,72], and suppressed the expression of MMP-1 and MMP-3 at transcriptional and protein levels [33]. In another study, human embryonic lung fibroblast cells were treated with different concentrations of MRJPs (0.1–0.3 mg/mL) versus bovine serum albumin. MRJP-treated cells showed the highest proliferation activity, the lowest senescence, and the longest telomeres. Such effects were associated with upregulation of SOD1 and downregulation of mTOR, catenin beta like-1, and tumor protein p53 [39]. Apart from these limited in vitro studies, we could not locate any study that investigated the effect of RJ on lifespan in humans in vivo. However, several studies assessed the effect of RJ on promotion of wellbeing and prevention of severe diseases associated with increased early death—these studies may mirror the healthspan effects of RJ. Herein, we explore how RJ may support healthy aging in the general population.
It is becoming clear that certain metabolic pathways reduce longevity in humans by increasing the risk of serious illnesses that contribute to mortality e.g., diabetes mellitus, metabolic syndrome, cardiovascular diseases, and cancer [73]. The life-expanding effect of RJ possibly originates from its antioxidant and anti-inflammatory properties, which can promote healthy aging by improving glycemic status, lipid profiles, and oxidative stress—and hence can prevent the occurrence of various debilitating metabolic diseases [13,14]. In accordance, administration of RJ in healthy volunteers was associated with improved indicators of physical wellbeing (erythropoiesis and glucose tolerance) [74].
.../...
F O R T H E R E S T O F T H E S T U D Y, P L E A S E V I S I T T H E S O U R C E .
.